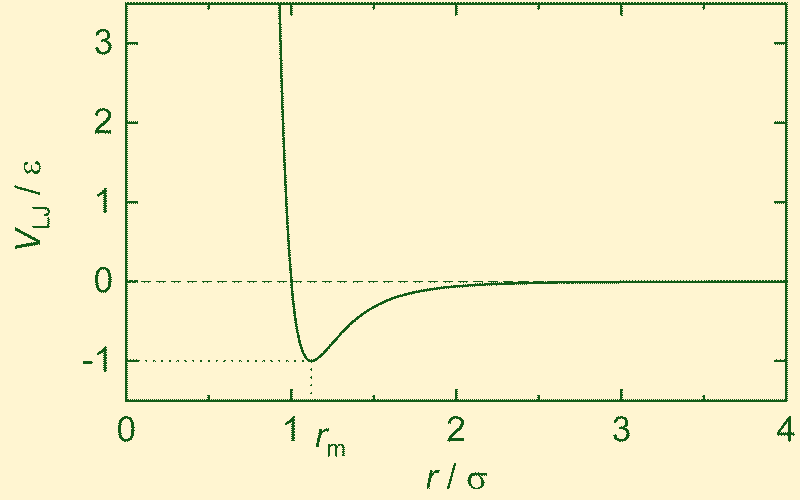
The Lennard–Jones potential is defined as:
\[ V(r) = 4\epsilon \left[ \left(\frac{\sigma}{r}\right)^{12} - \left(\frac{\sigma}{r}\right)^6 \right] \]where:
- \(r\) is the distance between two interacting particles,
- \(\sigma\) is the characteristic interaction distance,
- \(\epsilon\) is the depth of the potential well.
The first term,
\[ \left(\frac{\sigma}{r}\right)^{12}, \]represents strong short-range repulsion. As particles approach one another, the energy rises rapidly, preventing them from occupying the same space.
The second term,
\[ -\left(\frac{\sigma}{r}\right)^6, \]represents attraction. At moderate distances, this attractive force lowers the energy of the system and encourages association.
The balance between these opposing tendencies produces a stable equilibrium at:
\[ r_{\text{min}} = 2^{1/6}\sigma \]where the potential energy reaches its minimum value:
\[ V(r_{\text{min}}) = -\epsilon \]Stable forms arise not from attraction alone, nor from repulsion alone, but from the dynamic balance between the two.
For protein quaternary structures, the total interaction energy may be expressed as:
\[ E_{\text{total}} = \sum_{i=1}^{N} \sum_{j=i+1}^{N} V_{ij}(r_{ij}) \]where \(V_{ij}\) represents the interaction between residues, atoms, surface patches, or learned structural features belonging to different protein subunits.
The native assembly can then be viewed as the configuration that minimizes the global energy:
\[ Q^{*} = \underset{Q}{\operatorname{argmin}} \; E_{\text{total}}(Q) \]In this view, quaternary structure is not imposed from above. It emerges from countless local interactions, each individually simple yet collectively capable of producing highly ordered architectures. The completed complex is therefore not merely a collection of proteins but a geometric solution to a multidimensional optimization problem shaped by physics, chemistry, and evolution.
The same mathematical principle that governs the interaction of particles can be seen at larger scales throughout nature: local constraints generate global order. Complex structures emerge because there exist configurations in which competing forces achieve balance.